南湖新闻网讯(通讯员 孙霄鹏)10月13日,作物遗传改良全国重点实验室、湖北洪山实验室代明球教授、李林教授课题组联合湖北洪山实验室、园艺植物生物学教育部重点实验室李峰教授课题组在《自然—生物技术》杂志(Nature Biotechnology)发表了题为“The role of transposon invertedrepeats in balancing drought tolerance and yield-related traits in maize”的研究长文,利用多组学和基因编辑策略,详细阐述了玉米产量和抗性平衡的遗传与分子机制,为玉米高抗高产精准分子设计育种奠定了理论基础,提供了优异基因资源。
玉米是我国种植面积最大、产量最高的粮食作物,对保障国家粮食安全至关重要。玉米是高大作物,一生需水旺盛,对干旱敏感;而我国玉米主栽区多位于干旱半干旱地区,每年因旱灾造成玉米大幅减产和巨大经济损失。
因此,培育抗旱玉米具有重要的现实意义。作物育种往往以追求产量为目标,将控制优异生产性状的基因组人为选择加以固定;而自然选择以物种的后代繁衍为目标,侧重于环境适应性基因组的选择与利用。近百年来,育种家将玉米产量大幅提高,但伴随而来的是玉米抗旱性的显著下降,表明产量性状和抗逆性状往往相互拮抗,且控制二者的遗传位点在基因组上紧密连锁。如何打破这种连锁,使产量和抗性达到最佳平衡是作物育种有重要应用价值的研究课题,而找到控制产量性状的干旱胁迫应答负调控位点并揭示其遗传与分子作用机制,是高抗高产作物育种面临的重大瓶颈科学问题。
小RNA(sRNA)是一类短片段的非编码RNA,在真核生物中广泛存在。sRNA,尤其是miRNA,被报道对植物的生长发育和逆境胁迫应答起到重要的调控作用。植物基因组上普遍存在一类反向重复结构(Inverted repeat, IR),能够产生大量的sRNA。尽管知道这些IR产生的sRNA能够在转录和转录后水平调控基因的表达,但IR对作物发育的具体调控功能直到近年才在大豆上被报道。作物产量和环境胁迫抗性都是由微效多基因控制的复杂农艺性状,涉及到全基因组范围内众多基因的表达及其调控。IR是否及如何通过产生sRNA调控基因的表达进而控制玉米产量和环境胁迫耐受性迄今未知。
通过高通量测序,作者获得了338份玉米关联群体在正常浇水(WW)和干旱胁迫(DS)生长条件下的sRNA表达组和其中197份群体的转录组。从中检测到三万多个响应干旱胁迫的sRNA并获得其群体水平的表达性状。sRNA对基因表达有调控作用。在群体范围内通过回归分析共发现6158对sRNA-基因调控关系,其中有许多已知的miRNA-基因调控关系(如miR168对AGO1c的调控、miR169 对NF-YA8的调控、miR167对ARF6的调控等),同时也发现许多新的sRNA对基因的调控关系。对上述sRNA表达性状进行全基因组关联分析(eGWAS),共获得6000来个调控sRNA表达的eQTL,其中有29个是eQTL热点,能同时调控20个以上sRNA的表达。比较分析调控sRNA表达的eQTL和调控基因表达的eQTL发现,有4722个eQTL能同时控制sRNA和基因的表达,表明sRNA和基因之间存在大量的全基因组协同调控关系。
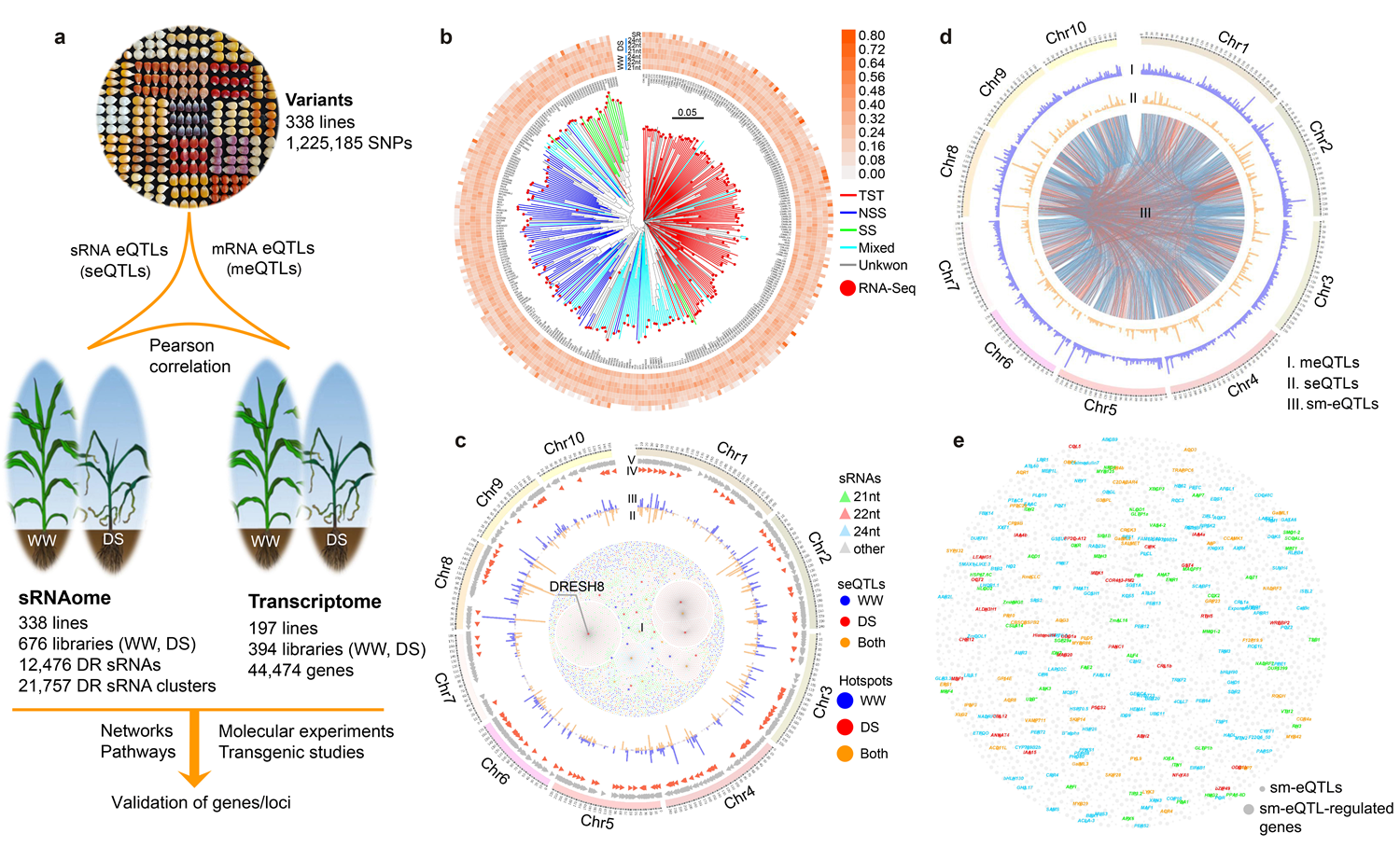
图1. 群体范围内鉴定sRNA和基因表达并解析其遗传调控机制
通过预测分析,发现29个eQTL热点所产生的sRNA调控2764个潜在的靶基因。其中,530个基因(19.2%)与植物繁殖相关,891个基因(32.2%)与胁迫应答相关,表明这些seQTL热点在调控玉米产量和环境适应性方面发挥重要功能。为了进一步解析作物产量和环境适应性分子调控机制,作者克隆了8号染色体上一个干旱特异性超级热点eQTLDRESH8,该热点eQTL与上千个sRNA表达相关,且仅在DS条件下被检测到。进一步分析发现DRESH8是一个由吉普赛类转座元件(TE)组成、有21.4kb的反向重复结构(TE-IR)变异。该TE-IR插入ZmPP2C16基因第三个内含子,并受到该基因启动子的调控,在干旱环境下诱导性启动表达。另外,DRESH8对插入的本地基因没有明显的调控作用,其可能通过产生sRNA在转录后水平对基因起调控作用。
群体中,缺乏DRESH8的玉米sRNA表达量低,但其干旱胁迫下的存活率高于携带DRESH8的玉米。进一步,通过基因编辑手段获得缺失DRESH8的转基因玉米,发现该转基因玉米比野生型对照玉米更抗旱,且其产生的sRNA比野生型显著减少,表明DRESH8是玉米群体内抗旱性变异和sRNA表达变异的因果变异位点。通过mRNA切割实验,发现DRESH8产生的sRNA对下游包括ZmMYBR38在内30个左右的基因有切割作用。通过在玉米和拟南芥中过表达ZmMYBR38基因来检测该基因的抗旱作用,发现两组过表达ZmMYBR38的转基因植物都比相应的野生型非转基因植物更耐旱,说明ZmMYBR38在植物抗旱中发挥保守和积极的作用。这些数据表明,DRESH8产生的siRNA通过介导下游靶标mRNA的切割来调控植物的耐旱性。
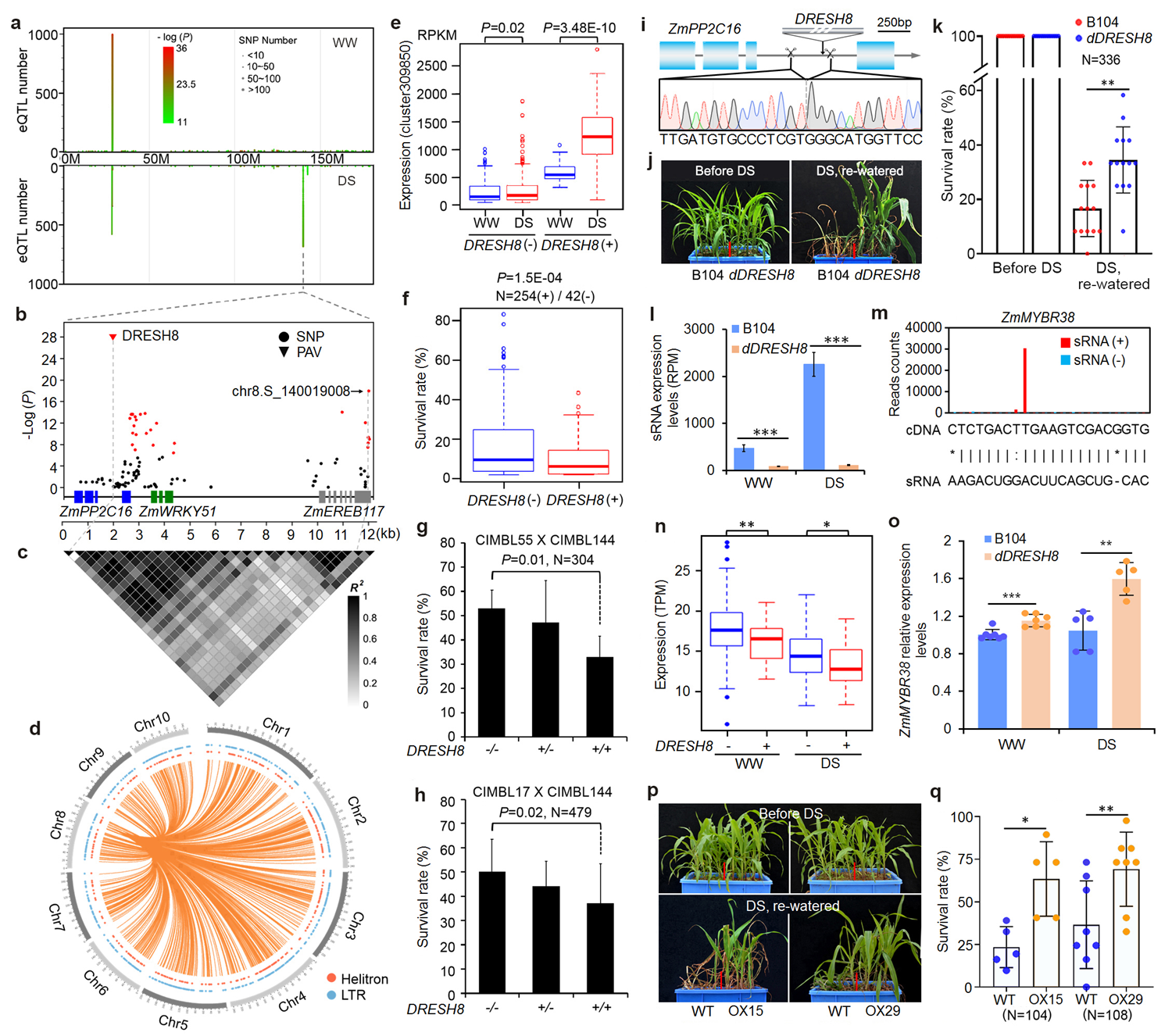
图2. DRESH8通过其产生的sRNA来调控靶标基因的表达进而抑制玉米抗旱
接下来作者详细研究了DRESH8的进化与选择。利用不同的分析手段均发现DRESH8位点存在较强的选择信号。利用包括玉米祖先大刍草、农家种和现代玉米自交系在内一共上千分玉米种质资源,进一步探究DRESH8的起源和传播。结果表明,DRESH8转座子的大致起源时间是51800年前,DRESH8的缺失可能是在较近的大刍草适应环境胁迫变化过程中被自然选择的,而DRESH8等位基因的存在则可能是在玉米的驯化和传播过程中被人为选择保留下来。考虑到DRESH8的存在不利玉米抗旱,是什么驱动了DRESH8被选择保留?为了回答这一重要问题,作者获取了两方面数据。一是降雨量与DRESH8的关系。他们发现,降雨量在种植含有DRESH8玉米的地区要比种植不含有DRESH8玉米的地区要高。二是产量与DRESH8的关系。他们发现,就产量性状而言(如穗长、穗直径、粒长、粒宽、粒厚、百粒重等),含有DRESH8的野生型玉米要比不含DRESH8的基因编辑转基因玉米提高5%-14%,即DRESH8的存在有利于提高玉米产量。基于这些结果,作者提出了一个模型来解释DRESH8位点是如何被选择的:在降雨充沛的环境中,农民可能选择种植有DRESH8的玉米以提高产量;而在干旱环境下,农民选择种植缺失DRESH8的玉米,这样可以去除DRESH8产生的sRNA对抗旱基因的抑制作用,从而提高玉米的抗旱性。以上这些数据表明,DRESH8能够介导耐旱性和产量性状之间的选择平衡,进而影响玉米在世界范围内的传播。
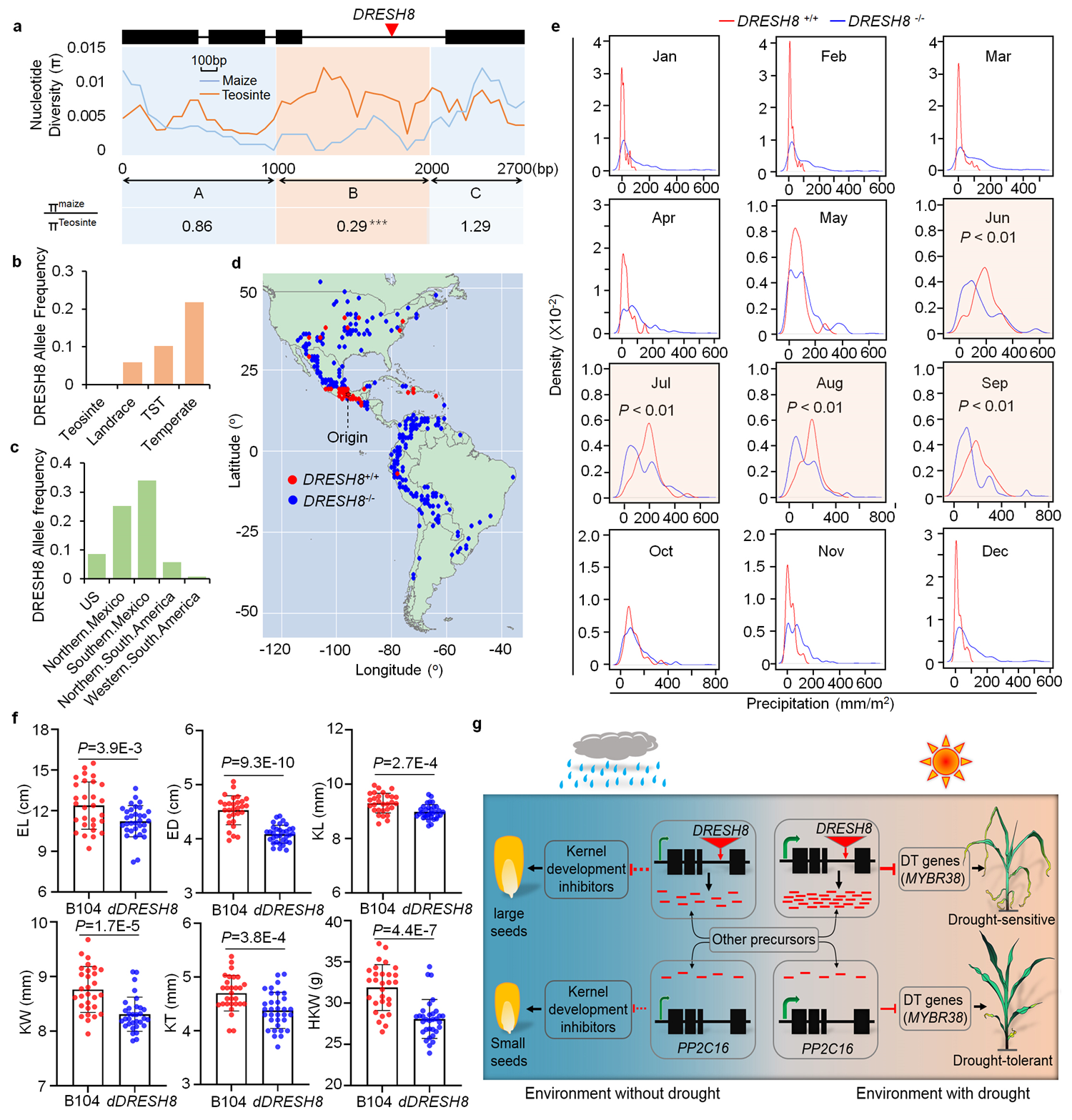
图3. DRESH8的进化与选择
为了更全面地描述IR在玉米响应干旱中的普遍作用,作者研究了全基因组范围内的IR结构。基于B73基因组,一共鉴定到8261个IR;尽管这些IR仅占玉米基因组的1.2%,却控制了42%左右的干旱应答sRNA的表达,表明IR在玉米干旱响应中发挥着广泛的作用。 玉米基因组85%都是TE,在8261个IR中,TE-IR占比高达90.7%,产生86.9%的IR sRNA。TE在形成IR结构后以产生22-nt sRNA为主,而不形成IR结构的TE则以产生24-nt sRNA为主。另外,由IR产生的22-ntsRNA占全基因组范围内22-nt sRNA的70%左右。在TE-IR中,大多数DNA转座子(除DHH和DTA外)形成较短的IR(10 kb)。IR产生的大多数21-和22-nt sRNA在干旱后上调表达,而且,这些IR sRNA还介导了包括低温和盐胁迫的应答反应,表明IR sRNA在调节植物响应不同环境胁迫方面具有普遍性作用。进一步发现,在驯化和改良的过程中,无论是IR长度还是数目都出现扩张,对粒长的选择可能是IR扩张的重要驱动因素之一。此外,有23个eQTL热点为TE-IR结构变异,其中有21个TE-IR产生的sRNA所调控的潜在靶基因富集到与产量和抗逆性相关的基因集中。综上所述,这些数据表明IR,特别是TE-IR,可能在调节玉米环境适应性与产量性状平衡上具有普遍作用。
作者发现了大量与干旱适应性和产量性状相关的环境特异性遗传调控因子。研究结果突出了TE在形成IR结构中发挥的关键作用。这些IR结构主要通过DCL2控制sRNA的表达,并参与转录后调控,IR的这种作用方式是玉米环境适应性和作物产量性状之间平衡的关键遗传与分子调控机制。这种由TE-IR结构驱动的平衡机制,为通过基因组精准设计来培育高抗高产作物铺平了道路。
华中农业大学作物遗传改良全国重点实验室博士后孙霄鹏、向艳丽博士(现根特大学博士后)、博士生豆楠楠、华中农业大学博士后张慧为论文的第一作者。作物遗传改良全国重点实验室和洪山实验室的代明球教授、李林教授以及园艺植物生物学教育部重点实验室的李峰教授为论文的通讯作者。作物遗传改良全国重点实验室和洪山实验室的严建兵教授也参与了该项工作。本研究得到了基金委国际合作和面上项目、十三五重点研发计划、中央高校基金项目、湖北洪山实验室重点研发项目的主要资助。
审核人 代明球
线上论文链接:https://www.nature.com/articles/s41587-022-01470-4
延伸阅读:【新闻特写】“抗旱?产量?鱼和熊掌可以兼得!”



 最近新闻
最近新闻